

Como as raízes absorvem nutrientes? Como os nutrientes são transportados para a planta? Entenda!
Entenda como as raízes absorvem nutrientes e como estes são transportados pela planta
Agrolink
- Anderson Wolf Machado

Índice dos conteúdos - clique para navegar
O que é a raiz? Qual sua função?
Anatomia das raízes
Como as raízes absorvem os fertilizantes / nutrientes?
Transporte de nutrientes das raízes para a parte aérea da planta
O que é a raiz? Qual sua função?
A raiz é o órgão especializado para a fixação da planta no solo e para a absorção de água e sais minerais em solução, podendo ainda desempenhar as funções de reserva de substâncias e de aeração em plantas aquáticas. A raiz é caracterizada como um órgão cilíndrico, aclorofilado que se distingue do caule por não se apresentar dividida em nós e internós e por não formar folhas ou gemas.
No interior da semente, o embrião consiste de um eixo hipocótilo-radicular, com um ou mais cotilédones na sua parte superior e na sua porção inferior está a radícula, ou primórdio do sistema radicular (Figura 1). Em muitas espécies a radícula não passa de um conjunto de células meristemáticas, enquanto que em outras, a radícula já se apresenta mais diferenciada.
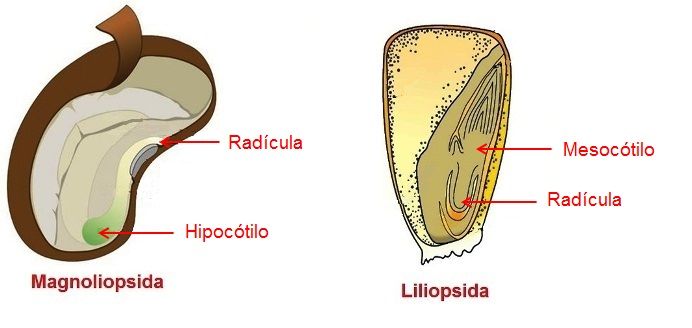
Figura 1 – Esquema mostrando o embrião em Magnoliopsida (Dicotiledôneas) e Liliopsida (Monocotiledôneas).
Quando a semente germina, a primeira estrutura a emergir é a radícula, que é responsável pela formação da primeira raiz da planta. Nas Gimnospermas e Magnoliopsida (Dicotiledôneas) esta raiz, geralmente, permanece toda a vida da planta e a partir dela se formam as raízes laterais. Nas Liliopsida (Monocotiledôneas), a raiz primária degenera-se precocemente e o sistema radicular que se desenvolve a seguir é formado por numerosas raízes adventícias, que se originam do hipocótilo, região caulinar acima da radícula. Em algumas Liliopsida, essas raízes adventícias iniciam o seu desenvolvimento ainda no próprio embrião.
Anatomia das raízes
Estrutura primária da raiz
A estrutura primária da raiz tem origem no meristema apical. A organização interna da raiz é bastante variada, mas é mais simples e mais primitiva do que a do caule. Os meristemas primários são os responsáveis pela diferenciação dos tecidos primários da raiz (Figura 2):
- A protoderme origina o revestimento primário da raiz - a epiderme;
- O meristema fundamental dá origem à região cortical, geralmente, formada apenas pelo parênquima;
- O procâmbio forma o cilindro vascular onde se encontra os tecidos vasculares primários.
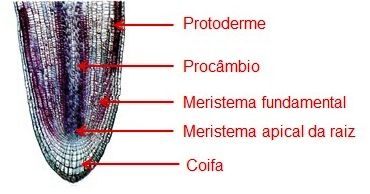
Figura 2 – Esquema mostrando a estrutura primária da raiz.
Existe uma nítida separação entre os três sistemas de tecidos que compõe a raiz (Figura 3):
a. Sistema dérmico – formado por epiderme;
b. Sistema fundamental – formado pelo córtex;
c. Sistema vascular – formado pelos tecidos vasculares (xilema e floema).
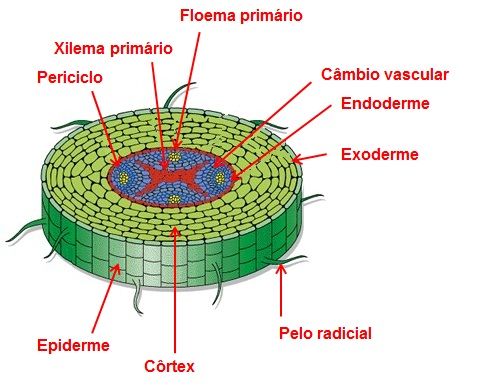
Figura 3 – Esquema mostrando os sistemas de tecidos da raiz.
Sistema dérmico - Epiderme
A epiderme da raiz é formada de células vivas, de paredes primárias e cutícula delgada (Figura 4). Nas espécies onde a epiderme persiste por mais tempo, as paredes de suas células podem apresentar maior cutinização ou até mesmo podem sofrer suberização.
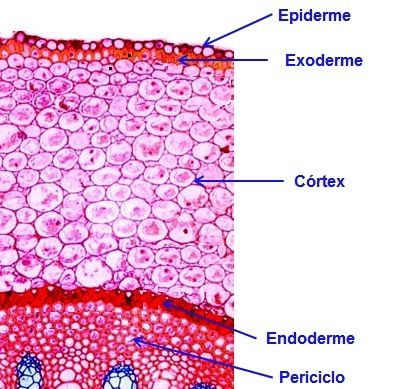
Figura 4 – Esquema mostrando os Sistemas dérmico e fundamental.
Nas raízes jovens, a epiderme especializa-se para a função de absorção e para isto desenvolvem numerosos pêlos radiculares (absorventes), que se formam a partir dos tricoblastos. A zona pilosa está restrita a uma faixa de poucos milímetros, não muito próxima do ápice da raiz. Nas regiões mais velhas esses pelos vão morrendo e sendo eliminados, enquanto novos pêlos radiculares vão sendo produzidos nas regiões mais jovens, mantendo assim a mesma extensão de zona pilosa (Figura 5).
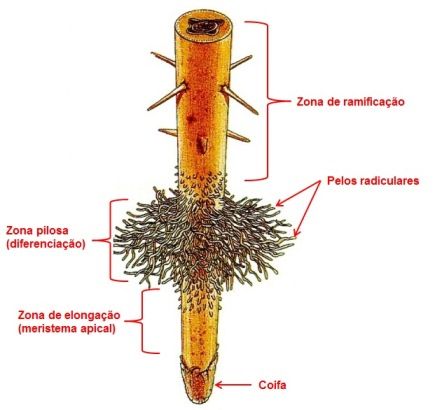
Figura 5 – Esquema mostrando zona pilosa.
Sistema fundamental - Córtex
O córtex da raiz é a região entre a epiderme e o cilindro vascular, e tem origem a partir do meristema fundamental. Esta região é formada por células parenquimáticas de paredes delgadas com numerosos espaços intercelulares. Nas plantas aquáticas ou naquelas que crescem em solos pantanosos pode haver a formação de um aerênquima na região cortical, para facilitar a aeração interna do órgão. O parênquima cortical da raiz é aclorofilado, exceto em raízes aéreas e em algumas espécies aquáticas.
A camada interna do córtex diferencia-se em uma endoderme e, frequentemente, as raízes desenvolvem uma ou mais camadas de células diferenciadas, na periferia do córtex, que se forma logo abaixo da epiderme denominada de exoderme.
ENDODERME
Na região de absorção da raiz primária, as paredes das células da endoderme apresentam um espessamento de suberina, em forma de fita, completamente impermeável, que é denominado estria ou faixa de Caspary (Figura 6).
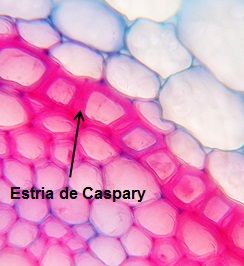
Figura 6 – Observação da região da endoderme apresentando a estria de Caspary (aumento 1000X).
Esta faixa suberizada é formada durante a diferenciação da célula endodérmica e faz parte da parede primária das células. A deposição da suberina é contínua desde a lamela média e, nesta região, a membrana plasmática também se encontra fortemente ligada com essas estrias. Desta forma, existe uma região de forte adesão entre o protoplasma das células endodérmicas vizinhas.
Como as células da endoderme estão perfeitamente justapostas e ainda se encontram fortemente ligadas umas às outras pelas estrias de Caspary, fica assegurado que somente as substâncias que passam pela seletividade da membrana plasmática, cheguem ao xilema, e daí sejam conduzidas para o restante do corpo da planta.
Nas espécies, que apresentam crescimento secundário como as Magnoliopsida (Dicotiledôneas) e as Gimnospermas lenhosas, as células endodérmicas não desenvolvem nenhum outro tipo de espessamento além das estrias de Caspary e, eventualmente, são eliminadas junto com o córtex durante o crescimento secundário.
No entanto, nas raízes que não apresentam crescimento secundário, especialmente entre as Liliopsida (Monocotiledôneas), a endoderme permanece e apresenta modificações de parede. Nas regiões mais velhas destas raízes, acima da região de absorção, as paredes das células endodérmicas vão sendo recobertas por uma lamela de suberina ou endodermina e num terceiro estágio, são recobertas com uma espessa camada de celulose lignificada. Esse espessamento secundário pode se dar de modo uniforme em todas as paredes da célula ou ser irregular, mais fino, ou mesmo ausente, o que leva as células endodérmicas a adquirirem o aspecto de U.
Além disso, nestas raízes, as células da endoderme em frente aos elementos do protoxilema não desenvolvem esses espessamentos secundários, sendo denominadas de "células de passagem", pois permitem a passagem de água e sais através da membrana plasmática (Figura 7).
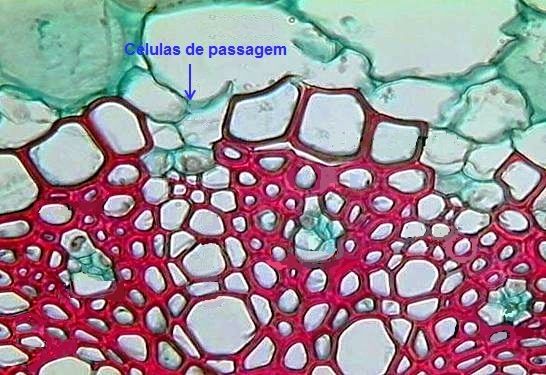
Figura 7 – Detalhe do cilindro vascular mostrando as células de passagem.
EXODERME
Algumas raízes apresentam uma ou mais camadas de células logo abaixo da epiderme ou do velame, diferentes das demais células da região cortical, formando uma exoderme. Frequentemente, a exoderme apresenta estrias de Caspary como a endoderme, mas comumente, as suas células apresentam uma camada de suberina recobrindo a parede celular.
Cilindro vascular
O cilindro vascular da raiz diferencia-se a partir do procâmbio e é formado por uma ou mais camadas de tecido não vascular chamado de periciclo. O periciclo é a camada mais externa do cilindro vascular, localizando-se logo abaixo da endoderme. Nas raízes jovens é constituído de parênquima, cujas paredes das células são celulósicas e delgadas. Nas angiospermas e gimnospermas, o periciclo está relacionado com atividades meristemáticas. As raízes laterais são formadas a partir do periciclo.
Nas raízes que apresentam crescimento secundário, o felogênio e parte do câmbio vascular também se originam no periciclo. Nas monocotiledôneas que não apresentam crescimento secundário, frequentemente, o periciclo torna-se lignificado nas regiões mais velhas da raiz.
Na raiz os tecidos vasculares não formam feixes vasculares, uma vez que o xilema primário forma um maciço central, provido de projeções que se dirigem para a periferia do cilindro vascular, e o floema forma cordões que se alternam com as arestas do xilema, distribuídos próximos da periferia do cilindro vascular, internamente ao periciclo. Quando o xilema não ocupa todo o centro do cilindro vascular, forma-se uma medula parenquimática nesta região central, que pode vir a esclerificar-se nas regiões mais velhas da raiz (Figura 8).
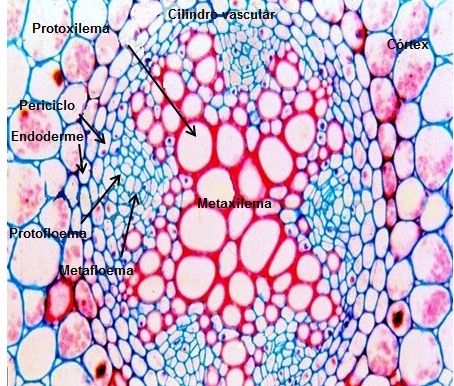
Figura 8 – Esquema mostrando o cilindro vascular da raiz.
Estrutura secundária da raiz
O crescimento secundário da raiz consiste na formação de tecidos vasculares a partir do câmbio vascular e da periderme. O início do câmbio vascular dá-se a partir de divisões de células do procâmbio (Figura 9) que permanecem indiferenciadas entre o floema e o xilema primários. Em seguida, as células do periciclo localizadas em frente aos pólos de protoxilema, começam a se dividir e, finalmente, o câmbio circunda todo o xilema. Neste estágio o câmbio apresenta o formato oval nas raízes diarcas; estrela de três pontas nas raízes triarca, e estrela de quatro pontas nas raízes tetrarcas.
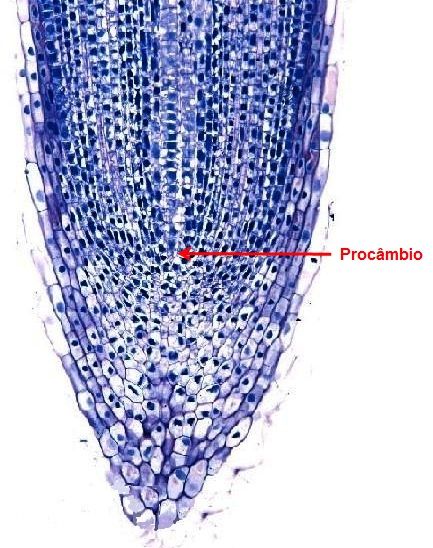
Figura 9 – Esquema mostrando o procâmbio.
Em seguida, as células do periciclo localizadas em frente aos pólos de protoxilema, começam a se dividir e, finalmente, o câmbio circunda todo o xilema. Neste estágio o câmbio apresenta o formato oval nas raízes diarcas, estrela de três pontas nas raízes triarca e estrela de quatro pontas nas raízes tetrarcas.
As regiões do câmbio localizadas em frente ao floema entram em atividade antes daquelas regiões do câmbio derivadas do periciclo. Com a formação do xilema secundário, o câmbio destas regiões, vai sendo deslocado para fora, até adquirir um contorno circular. A adição de tecidos vasculares secundários vai provocar um aumento no diâmetro do cilindro vascular e esse desenvolvimento leva a região cortical da raiz a apresentar modificações, para acompanhar esse crescimento.
Embora a maioria das raízes apresente crescimento secundário da maneira descrita aqui, inúmeras variações deste crescimento podem ser encontradas em várias espécies. Por outro lado, as raízes da maioria das monocotiledôneas geralmente não apresentam crescimento secundário.
COIFA
A coifa é uma estrutura que reveste o ápice radicular protegendo o meristema apical. A coifa tem origem no caliptrogênio, que é parte do meristema apical da raiz e é formada de células parenquimáticas, vivas, que secretam mucilagens que auxiliam na proteção do ápice radicular e na penetração da raiz através do solo (Figura 10).
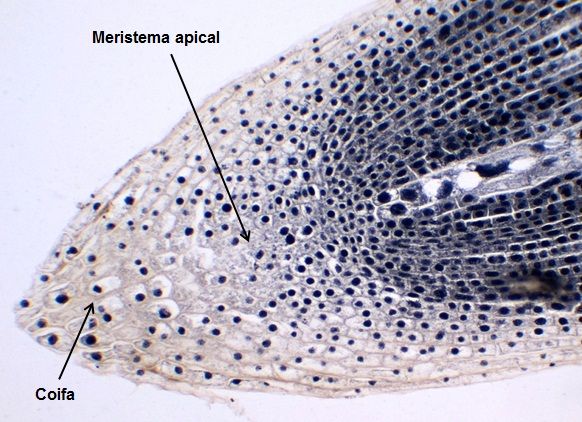
Figura 10 – Esquema mostrando a Coifa e o Meristema apical.
À medida que as raízes crescem e penetram no solo, as células da periferia da coifa vão sendo eliminadas e novas células vão sendo continuamente produzidas. As espécies aquáticas, geralmente, apresentam coifas bem desenvolvidas para proteger o meristema apical do ataque de microrganismos, abundantes do meio aquático. O direcionamento do crescimento geotrópico da raiz se dá em resposta à gravidade.
Como as raízes absorvem os fertilizantes / nutrientes?
Durante muito tempo pensou-se que os elementos (nutrientes) contidos na solução do solo se movimentassem em favor de um gradiente de concentração, indo de um local de maior concentração (exterior da planta) para um de menor concentração (interior da planta), movimento este conhecido por difusão. Porém, após comparações entre o suco celular e o meio, verificou-se que a concentração dos elementos no meio interno (suco celular) era muito maior do que o meio externo (solução do solo), havendo, também, seletividade na absorção dos íons dos nutrientes.
A absorção iônica é caracterizada por:
- Seletividade – certos elementos minerais são absorvidos preferencialmente;
- Acumulação – a concentração dos elementos, de modo geral, é muito maior no suco celular do que na solução externa;
- Genótipo – existem diferenças entre espécies de plantas nas características de absorção.
Sistema solo/planta
Os solos são compostos por três fases: sólida, líquida e gasosa. A concentração de nutrientes na solução do solo depende muito do teor de água no solo, do pH, da atividade microbiológica e da aplicação de fertilizantes.
Na fase sólida, a fração argila, além de influenciar várias propriedades físicas e físico-químicas do solo, determina a capacidade de troca iônica do solo. Já na fase líquida, a solução do solo é o compartimento de onde a raiz retira ou absorve os elementos essenciais. É constituída de uma solução de eletrólitos em equilíbrio com a fase sólida, que é o reservatório do solo.
Absorção de nutrientes pelas raízes
Para que os nutrientes sejam absorvidos é necessário que ocorra o contato íon/raiz. A capacidade das plantas para obter água e nutrientes minerais do solo depende da sua capacidade para desenvolver um extensivo sistema radicular. O desenvolvimento do sistema radicular de mono e de dicotiledôneas depende, em grande parte, da atividade do meristema apical das raízes. A absorção de íons é mais pronunciada em raízes jovens. Além disso, a taxa de absorção de íons das raízes diminui na região mais distante do ápice radicular. No entanto, esta tendência varia bastante, dependendo de fatores, como tipo de íon (nutriente), estado nutricional e espécie vegetal.
O encontro dos íons com a raiz pode envolver três processos diferentes: absorção por difusão, absorção por fluxo de massa e absorção por interceptação radicular.
Absorção de nutrientes por difusão
Na difusão, nutrientes minerais movem-se de uma região de maior para outra de menor concentração. A absorção de nutrientes pela raiz diminui a concentração dos íons nesta região e favorece a difusão em direção à superfície radicular. Quando a difusão é lenta, cria-se uma zona de esgotamento do nutriente próximo à superfície da raiz. Elementos que tendem a ficar imobilizados, como o fósforo, que possuem dificuldade de ser carreados por fluxo de massa, chegam às raízes por difusão. Além disso, o tamanho do sistema radicular é muito importante para a absorção de elementos que entram em contato com a raiz por difusão.
Absorção de nutrientes por fluxo de massa
Ocorre quando a água absorvida pelas plantas flui ao longo de um gradiente de potencial hídrico, arrastando consigo os nutrientes dissolvidos na solução do solo para próximo da superfície radicular onde ficam disponíveis para a absorção. Os nutrientes muito móveis na solução do solo, como o nitrato e o cálcio, tendem a chegar até as raízes por fluxo de massa.
O fluxo em massa ocorre por diferença de pressão, a qual é determinada, primariamente, pela taxa de transpiração. Assim, a quantidade de nutriente suprida por fluxo em massa depende da transpiração e da concentração do nutriente na solução do solo.
Absorção de nutrientes por interceptação radicular
Ocorre quando as raízes, ao crescerem explorando o solo em todas as direções, entram em contato direto com os nutrientes a serem absorvidos.
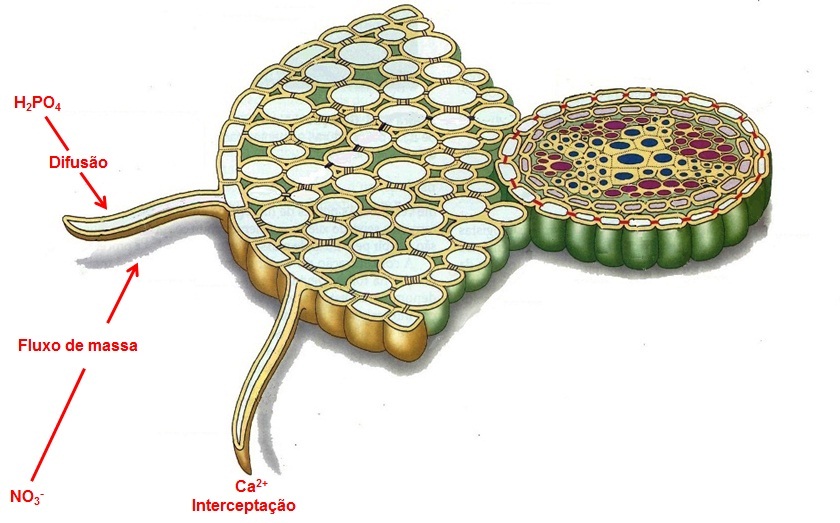
Figura 11 – Esquema mostrando os tipos de contato íon/raiz.
Absorção de nutrientes por micorrizas (Fungos micorrízicos arbusculares - FMA)
As raízes da maioria dos vegetais apresentam uma associação peculiar com determinados fungos, que desempenham um importante papel na nutrição destas espécies. Tais associações são denominadas micorrizas (fungo-planta). Uma micorriza é uma associação simbiótica entre um fungo não patogênico e as células de raízes jovens, particularmente as células epidérmicas e corticais.
Apesar de invadir a região cortical, o fungo não afeta o desenvolvimento da raiz. As hifas do fungo crescem também para o meio externo (solo), aumentando bastante a capacidade para absorver alguns nutrientes encontrados em baixas concentrações na solução do solo, como fosfato e alguns micronutrientes (Zn, Cu). Por sua vez, as raízes estariam secretando e fornecendo aos fungos, açúcares, aminoácidos e outras substâncias orgânicas necessárias ao seu desenvolvimento.
Trata-se de uma relação muito importante para a absorção de nutrientes de algumas espécies vegetais. Clique aqui para ler mais sobre as micorrizas!
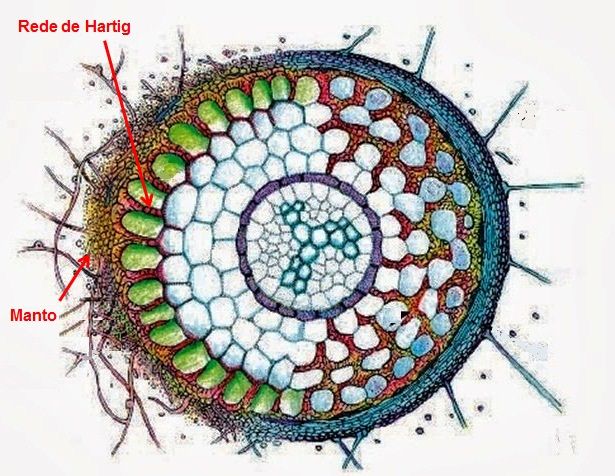
Figura 12 – Esquema mostrando raiz colonizada por ectomicorriza.
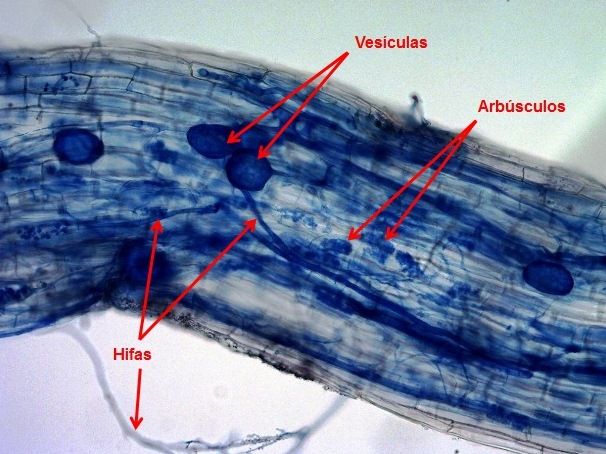
Figura 13 – Esquema mostrando as estruturas dos fungos micorrízicos arbusculares (hifas, vesículas e arbúsculos).
Transporte de nutrientes das raízes para a parte aérea da planta
Uma das características mais importantes das raízes é a sua capacidade de absorver e transportar íons do solo até o xilema e daí até a parte aérea. Ao chegar à superfície da raiz o íon pode seguir dois caminhos em direção ao xilema.
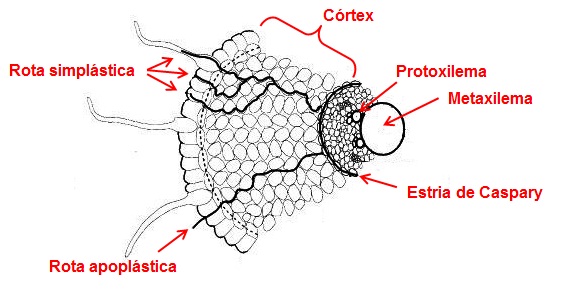
Figura 14 – Esquema mostrando rotas de transporte nos tecidos das raízes.
Rota apoplástica
O contínuo de paredes celulares e espaços intercelulares é conhecido como apoplasto e consiste em um sistema interconectado que engloba as células da epiderme e do córtex, em que a água e os íons inorgânicos movimentam-se livremente sem nenhuma restrição. A penetração de solutos neste espaço é rápida e, não é afetada por baixas temperaturas ou por inibidores metabólicos, sendo, portanto, inteiramente passiva e não envolvendo nenhuma membrana biológica. O apoplasto constitui, em média, de 10 a 15% do volume total das raízes.
Rota simplástica
Os citoplasmas de células vizinhas, conectadas através dos plasmodesmas, formam um contínuo, que é conhecido como simplasma ou simplasto, por onde os íons e moléculas podem também se mover. Isto ocorre em função das inúmeras plasmodesmas interligando as diversas células.
Entre o córtex e o cilindro central existe uma camada de células especializadas, a endoderme. Nessa camada de células se formam as estrias de Caspary, que bloqueiam efetivamente a entrada de água e de íons minerais no cilindro central, via apoplasto.
Desta forma o movimento do íon:
- Pode entrar via simplasto imediatamente na membrana plasmática das células epidérmicas (inclusive nos pêlos radiculares) ou ele pode se difundir entre as células da epiderme e córtex, via apoplasto;
- Do apoplasto do córtex, um íon pode difundir-se radialmente para a endoderme ou entrar via membrana da célula cortical, no simplasto;
- Em todos os casos, o íon deve entrar no simplasto, antes que ele chegue ao cilindro central, devido a presença das estrias de Caspary nas células da endoderme.
Após o íon ter entrado no cilindro central através do simplasto, ele continua a se difundir de célula para célula. Finalmente, o íon retorna para o apoplasto (do cilindro central) e difunde-se para dentro do xilema. Novamente, as estrias de Caspary evitam que o íon retorne para o apoplasto do córtex. Assim, a planta pode manter uma maior concentração iônica no xilema do que no meio em que a raiz está crescendo (solução do solo).
Os nutrientes minerais, uma vez no xilema, são carreados para a parte aérea pelo fluxo transpiratório. Algumas vezes, a ascensão da seiva no floema é promovida pela pressão radicular, quando os solos estão úmidos e a umidade relativa do ar é alta, tal como ocorre durante as primeiras horas do dia (transpiração praticamente ausente).
Na parte aérea, alguns nutrientes minerais podem ser redistribuídos pelo floema, particularmente, os que são muito móveis.
Por fim, recomendamos o vídeo abaixo onde o Engenheiro Agrônomo João Vitor Pasquetto exemplifica a importância de uma raiz bem desenvolvida quanto à captação de água.
José Luis da Silva Nunes - Eng. Agrº, Dr. em Fitotecnia

